Last week, I wrote about my misgivings over the 5 degrees of separation (actually, of misalignment) between the rotatable polarizer’s positioning on the microscope stage and the polarization of the microscope’s laser. In discussing with Rudolf Oldenbourg (MBL), I learned that the misalignment does not matter. What does matter is that the peaks and valleys are every 90º and so indeed they are. The five degrees of skew will add a baseline but that is easily removed. Even better, when I tried to reproduce the skew this past week, it had vanished like morning mist. This problem is as red as a herring ever gets. And I’ll take it.
Feeling that my calibrations were, if not perfect, at least not terrible, I decided to look at some celery xylem. At the end of last year, I wrote about isolating xylem cells from celery stalks to use as a standard. They xylem is easy to dissect out of the stem and the cell thickenings (aka ribs) are contain highly aligned cellulose running parallel to the rib. The orientation of the rib is easy to see in the microscope and so the calculated fluorescence anisotropy should be found to aligned in the direction of the ribs.
I imaged a few celery xylem cells stained with Congo red. I was underwhelmed. The ribs were certainly fluorescent but they were modestly anisotropic. Modesty is generally considered a virtue, but in this case, not so much.
Then I imaged unstained xylem. This is material prepared identically except for no incubation in Congo red. This xylem was not only fluorescent, the fluorescence was anisotropic (Figure 1).
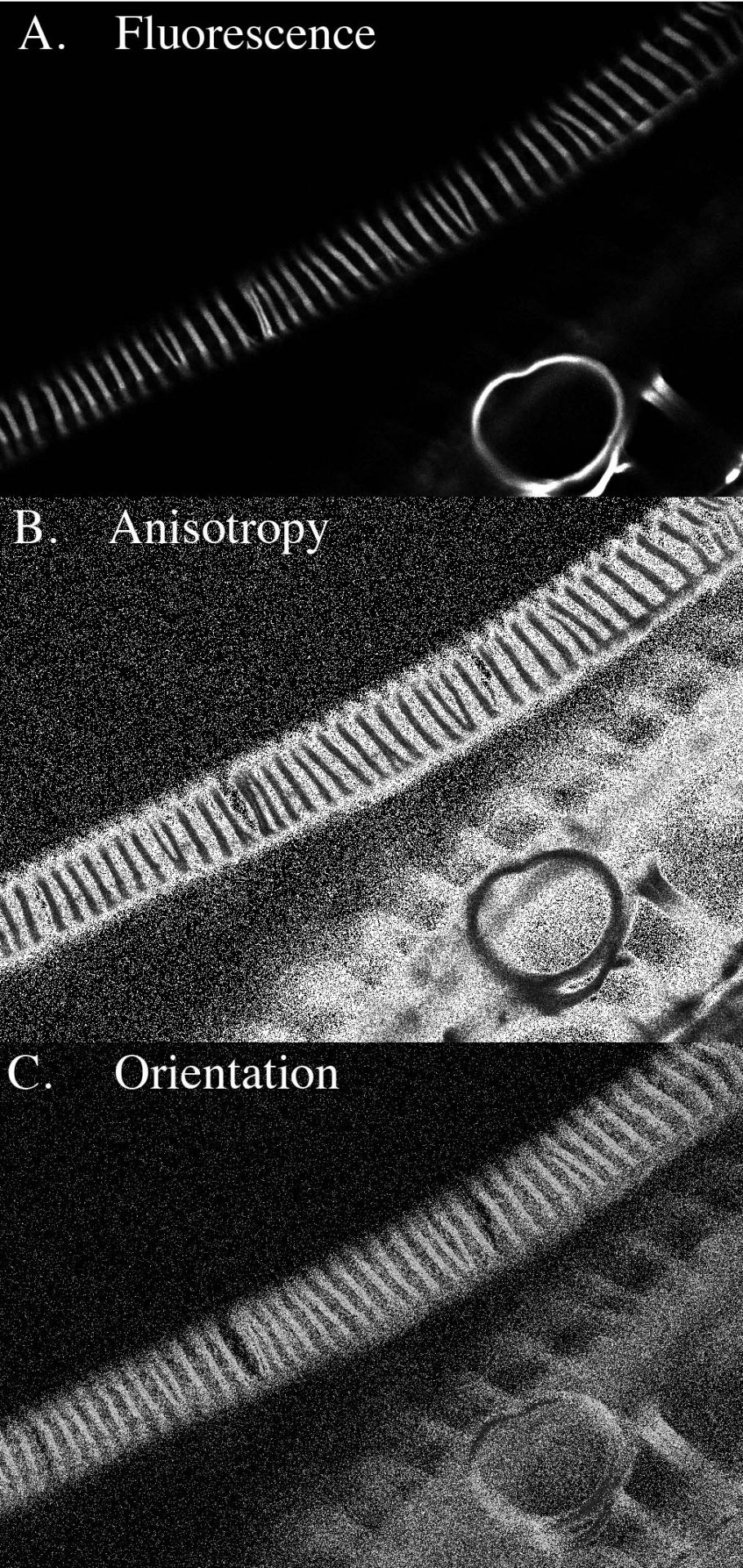
Figure 1. Unstained celery xylem. A. Standard fluorescence image. The bright, short lines running across the image from lower left to upper right and looking like railroad ties are the xylem thickenings. The ribs actually form a gently-sloped helix around the sides of the cell, excluded from the image here because of the narrow focal plane of confocal imaging. At lower right, there is a ring, a thickening that was pulled out from the cell by the dissection. B. Anisotropy image. Here, the brightness of each pixel is proportional to the amount of fluorescence anisotropy; that is, the brighter the pixel the more aligned are the fluorescent molecules. While the region between the ribs is very bright, there is little absolute fluorescence in this region (see A). The ribs themselves have substantial anisotropy (they are brighter than black). C. Orientation image. Here, the brightness of each pixel is proportional to the direction of net alignment of the fluorescent molecules. The uniform intensity of the ribs is consistent with a uniform orientation of fluorescent molecules; the absolute value of the intensity (the gray level) is consistent with the molecules being oriented, more or less, parallel to the ribs. In the ring at lower right, the intensity changes as approrpriate for the circular symmetry.
No, I didn’t screw up and image a stained slide by mistake. I am confident of this not only because the slides with stained xylem are visibly pink when handled in the lab. This slide is clear as water.
That unstained xylem fluoresces is not surprising. Fluorescence in a sample without added stain is termed auto-fluorescence; xylem is well known to be auto-fluorescent. The reason is lignin. Lignin is a kind of caulk that the plant smears through its mature cell walls to make them stronger and impervious to attack by pathogens. Xylem is typically highly lignified. Maybe, this is the first (world premier?!) observation that the auto-fluorescence in xylem thickenings is anisotropic. That’s possible. But again not too surprising in view of the underlying cellulose being so well aligned. Lignin assembling in, on, and around the cellulose is likely to become structured in conformity.
But for my project, there are two potential consequences to the celery xylem’s auto-fluorescence. First, impregnation with lignin might reduce the ability of dyes (e.g., Congo red) to bind, thereby making the signal weaker. Second, the fluorescent lignin molecules might interact with the added fluorescent stain and cause some anomalous behavior. The first is just annoying, the second might render celery xylem as a standard … problematic.
As a workaround, I decided to try imaging at a longer wavelength, in this case 561 nm, because auto-fluorescence from lignin goes down as the wavelength gets longer. My choice is ironic; I publicly bemoaned starting to work at this wavelength because it is not optimal for either Congo red or fast scarlet. Turns out, I might have been right after all.
I made a new calibration (gnash!) for 561 nm and found out that indeed, the auto-fluorescence, w while not zero, is much reduced. I needed to really crank the laser to see anything.
This coming week, I need to refine the 561 nm calibration and see how stained xylem is handled. Presumably, any interaction will be minimized by keeping the laser power well below that needed to excite auto-fluorescence. And I also need to take seriously the problem of the background.